为重症救治赋能
为患者康复加速

登录
注册
登录方式
方式一:
PC端网页:www.rccrc.cn
输入账号密码登录,可将此网址收藏并保存密码方便下次登录
方式二:
手机端网页:www.rccrc.cn
输入账号密码登录,可将此网址添加至手机桌面并保存密码方便下次登录
方式三:
【重症肺言】微信公众号
输入账号密码登录
注:账号具有唯一性,即同一个账号不能在两个地方同时登录。
王湘芸 上海和睦家医院长宁院区呼吸内科 发布于2025-09-01 浏览 1020
 收藏
收藏
作者:王湘芸
单位:上海和睦家医院长宁院区呼吸内科
急性呼吸窘迫综合征(ARDS)作为重症监护病房(ICU)常见的危重症,其高发病率与高死亡率一直是临床治疗的难题。近年来,随着微生态研究的深入,呼吸道与肠道微生态失衡在ARDS发生发展中的作用逐渐凸显,“肠-肺轴”的双向调控机制为理解和治疗ARDS提供了全新视角。本文将系统梳理呼吸道/肠道微生态与ARDS的临床关联、肠-肺轴的作用机制、微生态失衡的诊断价值、靶向治疗的临床转化及未来挑战,为临床实践与科研探索提供参考。
1. ARDS的未满足需求
ARDS年发病超300万,占ICU患者的10%。尽管肺保护通气策略(低潮气量+PEEP)使死亡率下降10%,但仍有46.1%的患者死于多器官功能障碍综合征(MODS)。这一数据揭示了单纯呼吸支持无法阻断全身炎症级联反应的核心矛盾——当前缺乏针对炎症风暴、上皮损伤等失调生物途径的有效药物,亟需从微生态角度寻找新的干预靶点。
2. 肠道微生态的功能与全身影响
人体肠道内寄生着约1000种、数量高达100万亿的细菌、病毒、真菌等微生物,其中细菌占主导地位(占90%以上)。肠道微生态的核心生理功能包括维持肠道屏障、参与营养代谢、调节免疫及抵御病原体入侵。当肠道微生态失调(如菌群失衡)时,肠道屏障功能受损可引发局部或全身炎症,与感染、炎症性肠病等多种疾病相关。
临床研究证实了肠道微生态紊乱对全身的影响显著。2023年Le Guern等研究提示多重耐药菌的肠道定植可以加速肺炎病情的恶化。该研究发现耐碳青霉烯酶类肠杆菌科细菌(CRE)的肠道定植可加剧铜绿假单胞菌肺炎病情,使患者死亡风险增加1.79倍,并延长住院时间。通过粪便菌群移植可改善CRE肠道定植小鼠的肺部感染情况、死亡率以及免疫应答。2025年Kullberg等研究也揭示了厌氧肠道微生物群对脓毒症的保护作用。专性厌氧菌通过竞争营养、维护厌氧环境、产生短链脂肪酸丁酸盐以及增强免疫球蛋白的产生来提供对可能有害微生物的定植抗性。厌氧共生菌的丧失导致肠球菌和肠杆菌科的扩张,导致脓毒症预后不良。此外,2025年我国学者的研究发现,将ARDS合并抑郁患者的粪便移植给健康小鼠后,小鼠出现肺损伤、抑郁样行为,且体内嗜黏蛋白阿克曼菌和丙酸菌减少,小胶质细胞激活,揭示了ARDS相关抑郁症的微生物群依赖性机制,并为制定针对ARDS相关精神症状的新型预防策略提供了见解。
3. 呼吸道微生态的失调特征
呼吸道微生态失调后,核心菌群会发生变化——致病菌如假单胞菌、葡萄球菌富集增多,而健康菌群如普雷沃菌属、梭杆菌属消耗减少。呼吸道微生态的α多样性与机械通气时间呈负相关,通气时间越长,菌群多样性越低,表明微生态失调程度加深。
ARDS患者肺泡灌洗液(BALF)微生物中,85%源自咽喉部,5%来自肠道,且在ARDS发病早期就存在。随着病情进展,ARDS患者BALF中肠道菌群比例上升,这凸显了肠道菌群在ARDS发展中的潜在作用,早期BALF检测肠道菌科可以预测ARDS风险。2024年我国学者研究表明,PCCM住院患者肠道菌群中肠球菌科、乳杆菌科、红环菌科、梭菌科和坦纳菌科的数量显著增多。
4. 肺和肠道微生物群与宿主之间的相互作用
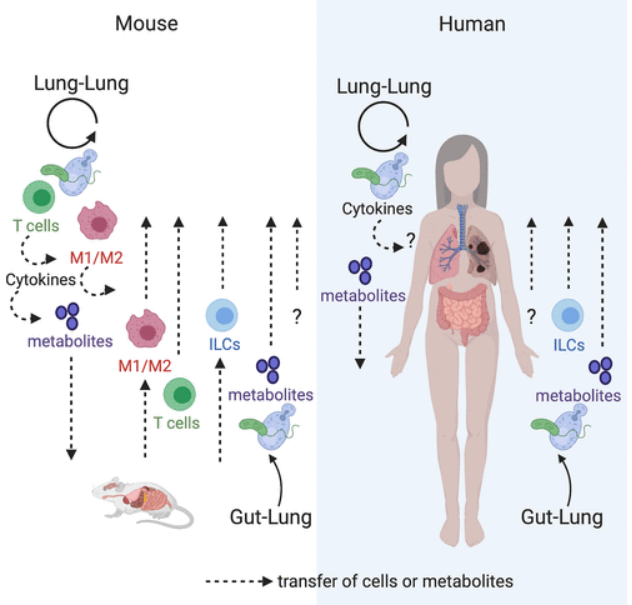
肠道微生物通过发酵膳食纤维产生短链脂肪酸(SCFAs),如乙酸、丙酸和丁酸,这些代谢产物不仅在肠道内维持屏障功能,还通过血液循环对远处器官发挥免疫调节作用。研究表明,SCFAs能够调控肺部局部的炎症反应,增强调节性T细胞的分化,进而降低过敏性炎症的发生风险。肠道菌群代谢的色氨酸衍生物和次级胆汁酸也被证明参与调节宿主免疫,进而影响肺部的免疫环境。参与的细胞包括T细胞、巨噬细胞极化状态(M1/M2)、先天淋巴细胞(ILC)、细胞因子和代谢物,包括脂多糖、小链脂肪酸、组胺、氧化脂质和前列腺素(图1)。
图1 肺和肠道微生物群与宿主之间的相互作用
图源:Am J Physiol Lung Cell Mol Physiol, 2020, 319(4):L710-L716.
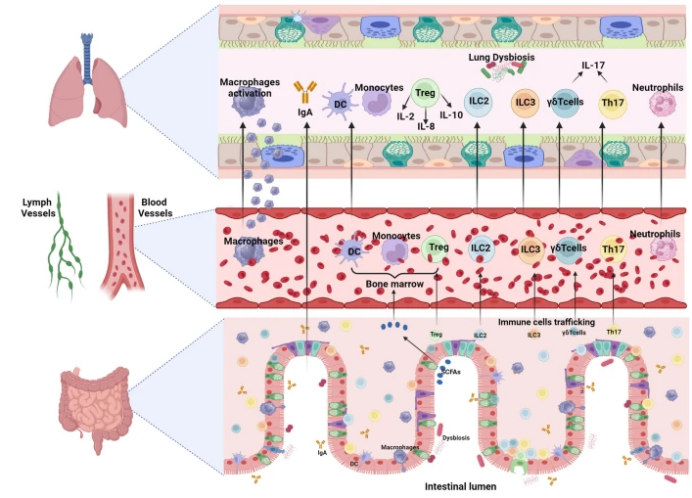
多项研究表明,肠道和肺之间存在特定的联系,统称为“肠-肺轴”。肠-肺轴进一步受到呼吸系统和消化系统微生物群、局部和远处免疫反应的影响。研究发现,133例肠易激综合征患者中有33%有呼吸问题,16%患有哮喘。慢性阻塞性肺疾病(慢阻肺)患者比健康对照组更有可能发展成溃疡性结肠炎或克罗恩病。粪便微生物移植手术和恢复肠道菌群已被证明可以改善败血症。这些研究均强调了保持健康的肠-肺轴通信在缓解肺部疾病方面的潜在益处。细胞从肠道迁移到肺部可能会影响呼吸道免疫,正如Tregs、Th17细胞、ILC2、ILC3和γδT细胞从肠道移动到肺部所示(图2)。
图2 细胞从肠道迁移到肺
图源:Crit Care, 2024, 28(1):220.
肠道和呼吸道上皮的功能存在差异,但两者都源自内胚层,由柱状上皮细胞构成,提供保护屏障。当肠道菌群失调时,屏障功能受损,出现“肠漏”现象,使内毒素(如脂多糖)和其他炎症介质进入血液,从而引发全身性炎症反应。系统性炎症进而可影响肺部微环境,诱发或加剧肺部疾病,如ARDS和肺纤维化。肠道菌群的破坏通过TLR4触发固有免疫反应,破坏肠屏障的完整性,增加细菌移位的风险。肠系膜淋巴亲水性和亲脂性成分可激活内皮细胞、中性粒细胞和单核细胞/巨噬细胞,诱导内皮细胞屏障功能障碍,延迟中性粒细胞凋亡,抑制红细胞变形性和树突状细胞功能,所有这些都会导致全身炎症播散和急性肺损伤。
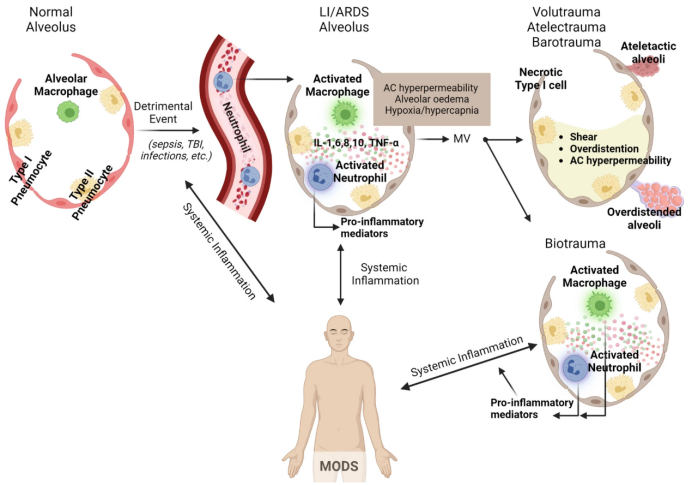
在肺损伤/ARDS患者中,肺毛细血管内皮发生损伤,导致活化的免疫细胞迁移到肺部,从而加剧肺部炎症反应(图3)。肺泡巨噬细胞局部分泌细胞因子以诱导趋化性并激活中性粒细胞,中性粒细胞进而释放各种促炎分子。使用机械通气可能会进一步加重肺损伤,导致肺泡过度扩张、肺泡反复周期性打开和关闭以及复杂炎症级联反应的启动,从而导致局部和全身炎症。这种炎症可以扩展到远处的器官和系统,加剧多器官功能障碍。
图3. 肺损伤/ARDS患者肺毛细血管内皮发生损伤,导致活化的免疫细胞迁移到肺部,从而加剧肺部炎症反应
图源:Crit Care, 2024, 28(1):179.
肠道在健康和危重疾病中的作用非常重要。在危重病患者中,维持体内平衡至关重要的肠道紧密连接蛋白完整性的改变会导致肠道完整性的丧失和通透性的增加。
在肺损伤/ARDS患者中,严重的急性炎症导致紧密连接蛋白的功能改变和肌球蛋白轻链激酶的激活,这与肠屏障功能障碍、黏液层完整性破坏、肠道炎症反应加剧和肠道生态失调有关。机械通气可能引起呼吸机相关性肺损伤(VILI),进一步加重全身炎症和肠道损伤。低潮气量机械通气期间的血流动力学改变和保护性机械通气诱导的高碳酸血症与心肌收缩性增加、全身血管阻力降低和血管张力改变相关,导致内脏血管张力和内脏微循环氧合的改变。肠道功能障碍和肠道完整性的丧失增强全身炎症,促进肠道细菌向肺部转移,肺生态失调进一步恶化。
在目前已明确的肠-肺轴相关机制中,肠道菌群引发肺部损伤的核心路径主要包括两个方面——细菌移位和炎症风暴。①屏障破坏与细菌移位:肠道屏障因TNF-α、IL-6等炎症因子上升而受损,闭锁蛋白、ZO-1等紧密连接蛋白减少,导致通透性增加,细菌趁机移位至肺部,引发炎症反应,这是肠-肺轴失调的关键环节。②代谢物失衡加剧炎症:短链脂肪酸(如丁酸)水平下降,使得组蛋白去乙酰化酶抑制失效,进而激活NF-κB,导致炎症失控。这种代谢物失衡在ARDS发生发展中起着推波助澜的作用,加剧了肺部炎症。免疫失调的后果就是组织进一步损伤。③免疫失调的级联效应:肠道细菌迟缓埃格特菌(E. lenta)在肠道中富集,增加了细菌性肺部感染的风险,它导致代谢物牛磺酸熊去氧胆酸(TUDCA)通过AMPK通路,引起血清循环到肺部的中性粒细胞功能障碍,加重组织损伤。
同样的,肺部损伤与机械通气也会产生反向影响:①VILI恶性循环:高潮气量通气导致肺泡牵张,还引发全身炎症反应,进而损伤肠道屏障。临床证据显示,机械通气患者血浆TNF-α水平上升与肠道通透性呈正相关,凸显了机械通气对肠道的不良影响。②高PEEP的内脏损伤风险:当PEEP超过15 cmH2O时,肝静脉氧饱和度下降,肠道缺血风险增加。这种高PEEP引起的内脏损伤会进一步扰乱肠道微生态,形成恶性循环,影响ARDS患者的预后。因此,建议ARDS患者中使用低潮气量和高水平PEEP,当使用高水平PEEP时,密切监测全身和局部灌注至关重要。
识别微生态失衡的标志物,对ARDS的早期预警、病情评估及预后判断具有重要意义。
1. 肠道代谢物标志物
(1)丁酸: 丁酸水平下降与肺损伤严重程度呈正相关, BALF中性粒细胞计数增加, 表明炎症反应加剧。丁酸作为短链脂肪酸的一种, 对维持肺部微生态平衡和抑制炎症具有重要作用。
(2)三甲胺-N-氧化物(TMAO):低水平TMAO与脓毒症患者撤机失败风险增加相关。TMAO是一种肠道代谢物, 其水平变化可反映肠道微生态状态, 对临床治疗决策具有指导意义。
2. 临床分型与预后模型
高炎症型ARDS患者标志物为IL-6、IL-8和血管生成素-2(ANG2)水平升高,死亡率较高,可能受益于靶向免疫调节治疗。通过多组学分析,可对ARDS患者进行精准分型,为个体化治疗提供依据。
联合代谢组(鞘氨醇-1-磷酸)和蛋白组(GAPDH、ENO1)的机器学习模型可以预测28 d死亡率,这种基于多组学数据的机器学习模型为ARDS预后评估提供了新的工具。
基于肠-肺轴机制的微生态靶向治疗已在临床转化中展现潜力,涵盖微生物调节、代谢干预、营养支持等多个维度。
1. 益生菌/益生元干预
(1)菌株特异性作用:双歧杆菌和乳酸菌可增强肠道屏障功能,使闭锁蛋白水平上升,同时抑制病原体定植。益生元(膳食纤维)可增加短链脂肪酸水平,促进Treg细胞扩增,抑制炎症反应。这些益生菌和益生元的作用为微生态靶向治疗提供了理论基础。
(2)临床使用规范:。益生菌和益生元的使用需注意细节,如用40℃以下的水冲服,避免与抗生素同服(间隔需≥2 h),以确保其疗效和安全性。
2. 代谢物补充与药物重定位
(1)丁酸盐补充:动物实验表明,丁酸盐可减轻LPS肺损伤,使中性粒细胞浸润减少50%,IL-6水平下降60%。丁酸盐作为一种短链脂肪酸,具有抗炎和修复屏障的作用,可作为ARDS治疗的潜在补充疗法。
(2)二甲双胍重定位:二甲双胍通过激活AMPK,可逆转TUDCA对中性粒细胞的抑制作用,在支气管扩张模型中表现出良好的治疗效果。这种药物重定位为ARDS治疗提供了新的思路和方法。
3. 营养与微生态调节
(1)n-3脂肪酸补充:ARDS患者使用n‑3脂肪酸(EPA、DHA)的随机对照试验荟萃分析显示:通常与其他生物活性营养素结合使用,如抗氧化剂,n‑3脂肪酸会减少机械通气时间和ICU住院时间,改善氧合,具有抗炎和调节免疫的作用,可改善ARDS患者的预后。
(2)维生素C的应用:CITRIS-ALI试验表明,维生素C可使脓毒症ARDS患者28 d病死率下降16%。对ARDS治疗具有重要意义。另一项单中心回顾性研究并未发现补充维生素B1和维生素C可改善需要有创机械通气的脓毒症ARDS患者的临床结果(病死率、呼吸机使用天数和住ICU天数)。
4. 呼吸支持优化
个体化调整PEEP。15 cmH2O的PEEP显著降低内脏灌注,建议在合并休克时将PEEP调整至小于10 cmH2O,并监测肝静脉氧饱和度。这种个体化的呼吸支持策略可减少对肠道等内脏器官的损伤,优化ARDS患者的治疗效果。
尽管微生态与ARDS的关联研究取得进展,但其临床转化仍面临诸多挑战。
1. 精准干预的瓶颈
(1)致病菌株机制不明:目前对致病菌和益生菌株的特异性机制尚不清楚,这限制了精准干预的实施。深入研究菌株机制对于开发针对性的微生态治疗方案至关重要。
(2)粪菌移植安全性数据缺乏:粪菌移植在ARDS治疗中的安全性数据不足,需要更多的临床研究来评估其风险和收益,为临床应用提供科学依据。
2. 技术突破点
(1)快速检测芯片:开发快速微生态检测芯片,可为临床医生提供及时的微生态信息,便于快速制定治疗方案。
(2)缓释纳米制剂:研发短链脂肪酸缓释纳米制剂,提高药物的稳定性和生物利用度,增强其治疗效果,为微生态靶向治疗提供更有效的工具。
3. 未来研究方向
(1)新型抑制剂研发:靶向TLR4/AMPK通路的新型抑制剂研发是未来研究的重点方向之一,有望为ARDS治疗提供更精准的干预手段。
(2)个体化营养配方:开发基于肠-肺轴导向的个体化营养配方,满足不同患者的需求,改善微生态状态,提高治疗效果。
(3)需从传统对症治疗(如肺保护性通气)转向更精准的干预策略:①核心目标:恢复肠-肺轴生态平衡;同步保护肺/肠道微生物组稳定性;②关键途径:调控肠道菌群组成,增强肠道屏障完整性;③预期效益:减少细菌/内毒素易位,降低ARDS发病率及严重程度,改善相关肠功能障碍。
呼吸道/肠道微生态与ARDS的关联为理解这一危重症提供了全新维度, “肠-肺轴”的双向调控机制揭示了全身微生态平衡对肺部健康的关键作用。当前研究已证实微生态失衡在ARDS发生发展中的核心地位,而靶向微生态的干预策略(如益生菌补充、代谢物调节、粪菌移植等)展现出临床转化潜力。未来,随着菌株机制研究的深入、检测技术的突破及个体化治疗的完善,微生态靶向治疗有望成为改善ARDS预后的重要手段,为重症患者带来新的希望。
作者介绍 王湘芸 上海和睦家医院长宁院区呼吸内科,副主任医师,中国医师协会呼吸医师分会危重症青委会委员,上海医学会呼吸分会慢阻肺学组成员,主要研究方向:肺癌、慢阻肺等疾病,发表多篇SCI和国内核心论著,主持并参与上海市、军队、医院多项课题,获医院首届优秀病例大赛一等奖、“十佳带教老师”称号,上海市内科和全科基地住培结业考官,PCCM国家专培结业考官。 声明: 本文仅用于学术内容的探讨和交流,不用于任何商业和推广,亦不作为最终的临床决策。临床实践需根据患者的具体情况选择适宜的处理措施。
 后可发表评论
后可发表评论




友情链接
联系我们
 公众号
公众号
 客服微信
客服微信